Chronic treatment with sildenafil stimulates Leydig cell and testosterone secretion
Correspondence: Dr Christina A. Peixoto Laboratório de Ultraestrutura Centro de Pesquisas Aggeu Magalhães FIOCRUZ Av. Moraes Rego s/n Recife CEP 50670-420 Brazil Tel.: (55) 81 210 12557 Fax: (55) 81 210 12516 E-mail: rb.zurcoif.maqpc@otoxiepc
Abstract
The phosphodiesterase type 5 (PDE5) inhibitor, Sildenafil, is a novel, oral treatment approach for pulmonary hypertension. As Leydig cells present PDE5, this study was conducted to investigate the effects of the chronic treatment with Sildenafil (25 mg/kg) on male Swiss Webster mice steroidogenesis. After a 4-week long experimental design, Leydig cells were analysed by morphological and immunocytochemical procedures. Serum testosterone was assayed by radioimmunoassay. Leydig cells presented noteworthy ultrastructural alterations, such as a vesicular smooth endoplasmic reticulum, large vacuoles scattered through the cytoplasm, enlarged mitochondria with discontinue cristaes and whorle membranes with vesicles at the periphery, which are typical characteristics of an activated steroid-secreting cell. Important immunocytochemical labelling for steroidogenic acute regulatory protein, cytochrome P450 side-chain cleavage enzyme and testosterone were detected in isolated Leydig cells. In addition, Sildenafil-treated mice showed significant increased levels of total testosterone. The results obtained in the present study are consistent with the hypothesis that the accumulation of cyclic guanosine monophosphate by PDE5 inhibition could be involved in the androgen biosynthesis stimulation. Important clinical implications of hormonal disorders should be taken into account for patients with pulmonary hypertension.
The cyclic nucleotide monophosphates (cAMP and cGMP) are important intracellular regulators of several processes, including smooth muscle motility, electrolyte homeostasis, neuroendocrine signals and retinal phototransduction. The labile gas, nitric oxide (NO), is a crucial mediator of smooth muscle relaxation and it interacts with soluble guanylate cyclases (sGC) in the cytoplasm to increase the conversion of guanosine triphosphate (GTP) into cyclic guanosine monophosphate (cGMP). On the other hand, natriuretic peptides activate membrane-bounded guanylate cyclases and accumulate intracellular cGMP. This increase in cGMP triggers a signal transduction cascade that encompasses the activation of cyclic nucleotide-dependent protein kinases A and G, subsequent phosphorilation of the actin-myosin system, as well as Ca 2+ channels and adenosine triphosphate-driven Ca 2+ pumps located in the outer cell membrane of sarcoplasmatic reticulum. This cascade leads to a reduction in cytosolic Ca 2+ and to smooth muscle relaxation (Uckert et al. 2006; Dimitriadis et al. 2008).
Sildenafil, a chemical compound designated as UK-92,480, is a water soluble citrate salt that was first synthesized by Pfizer in United Kingdom to treat hypertension and angina pectoris. Interestingly, this drug exhibited a different pharmacological effect, a marked penile erection, and became the first-line treatment option to erectile dysfunction (Kalsi & Kell 2004; Uthayathas et al. 2007). It has been reported that more than 20 million men worldwide are treated with this drug, and about $2 billion per year are spent on it (Sharma 2007).
The Sildenafil therapeutic possibilities arise from modulating intracellular levels of cGMP. This cyclic nucleotide is degraded into the inactive form by intracellular phosphodiesterase type 5 (PDE5) enzyme, which is present in the smooth muscle of the systemic vasculature and in platelets (Ghofrani et al. 2006). The main pharmacological action of Sildenafil is the inhibition of the cGMP-specific PDE5 with an inhibitory concentration (IC50) between 2 and 7 nM (Abbott et al. 2004), leading to cGMP accumulation and special effects in targeting organs.
Pulmonary hypertension is a rare and aggressive disease that affects children and adults with a bad prognostic and little life expectation. It can be classified as a primary or secondary hypertension, with various aetiologies. The cause of the primary pulmonary hypertension is not yet understood, but the secondary may be due to pulmonary, cardiac and extrathoracic malfunction (Uthayathas et al. 2007).
Sildenafil has revolutionized the treatment of pulmonary hypertension. Studies have demonstrated that this drug attenuated pulmonary hypertension by increasing the supply of blood to the lungs. This medicine reduced the right ventricular systolic pressure, right ventricular hypertrophy, the pulmonary artery muscularization and increased cGMP levels, suggesting that the NO-cGMP pathway contributed to the drug response (Zhao et al. 2001, 2003).
Several clinical research studies of pulmonary hypertension have been conducted using high dosages of Sildenafil for long time as additional therapy in children and adults (Schulze-Neick et al. 2003; Stocker et al. 2003; Karatza et al. 2004; Humpl et al. 2005; Preston et al. 2005; Ganière et al. 2006; Lobato et al. 2006). According to Karatza et al. (2005), Sildenafil therapy in series of children with pulmonary hypertension increased exercise capacity, and improved oxyhaemoglobin saturations without any side effects, appearing to be beneficial in the management of this disease.
However, important implications of the pulmonary hypertension treatment with high and diary doses of Sildenafil should be taken into account. Recently, PDE5 was localized by immunohistochemistry and western blotting in vascular smooth muscle cells, as well as in Leydig and peritubular cells of the rat testis (Scipioni et al. 2005). Therefore, it seems necessary to investigate whether compounds interfering with PDE5 activity, as Sildenafil, might affect testicular physiology.
The present work describes, for the first time, the effects of the chronic treatment with Sildenafil on mouse Leydig cells to contribute to a better knowledge about its action on the steroidogenesis.
Materials and methods
Chemicals
The Sildenafil was obtained from Pfizer Inc., New York, NY, USA; the anaesthetics Ketamine and Xylazine from Sespo Comércio e Indústria Ltda., Sao Paulo, Brazil; Hank’s balanced salt solution (HBSS) and Medium 199 (M199) from Gibco, Grand Island, NY, USA; Bovine serum albumin (BSA, fraction V) from Miles, Naperville, IL, USA; Percoll from GE Healthcare Bio-Sciences AB, Uppsala, Sweden; Collagenase (type I), Soybean trypsin inhibitor, Human chorionic gonadotropin (hCG), Leupeptin, Glutaraldehyde, Paraformaldehyde, Cacodylic acid, Sodium phosphate monobasic and dibasic heptahydrate, Osmium tetroxide, Calcium chloride, Potassium ferricyanide, Ammonium chloride, Tween 20, Lead (II) nitrate and Sodium citrate from Sigma Chemical Co., St Louis, MO, USA; SPIN-PON resin (Embed 812), LR-White resin and Uranyl acetate from Electron Microscopy Science, Washington, PA, USA; Acetone and Ethanol were purchased from Isofar Indústria e Comércio de Produtos Químicos Ltda., Rio de Janeiro, Brazil.
Experimental design
Forty pubertal Swiss Webster mice (obtained from the Centro de Pesquisas Aggeu Magalhães/FIOCRUZ, Recife, Brazil) aged 25 days and weighing 15–20 g were used in all experiments. Mice were examined for health status and acclimated to the laboratory environment at 25 °C and 12 h light:12 h dark photoperiod. The animals were housed in metal cages and fed a standard laboratory diet. The experimental group was composed of 20 animals that received 25 mg/kg body weight of Sildenafil for 4 weeks administered through the drinking water (Zhao et al. 2003). Body weights were recorded every day and the drug concentration in the water was adjusted to maintain the dose. The control group was also composed of 20 animals that received only pure water using the same procedure as described above. All experiments were carried out according to ethical guidelines (L-0035/08 – CEUA/FIOCRUZ).
After treatment with Sildenafil, the experimental and control animals were anaesthetized with ketamine (115 mg/kg, i.m.) and xylazine (10 mg/kg, i.m.), before blood collection by cardiac puncture without anticoagulant. Serum was separated and stored at –70 °C for radioimmunoassay of testosterone hormone. Testes were quickly dissected and fixed for morphological analysis.
Light microscopy
The testes were fixed in Bouin’s solution for 8 h. Then, they were dehydrated in an ethanol series and embedded in paraffin wax. Serial sections of 4 μm were cut using a microtome (Leica RM 2125RT), and stained with haematoxylin–eosin for histological observation (Weng et al. 2005).
Leydig cell isolation
Isolation and purification of mice Leydig cell-enriched preparations were modified from Hedger and Eddy (1986) and Wanderley and Negro-Vilar (1996). The testes were decapsulated with fine forceps without breaking seminiferous tubules, under aseptic conditions, and digested in an enzyme solution of 0.5 mg/ml collagenase, 0.2 mg/ml soybean trypsin inhibitor and 5 μg/ml leupeptin in Hank’s balanced salt containing 0.1% BSA (HBSS/BSA), pH 7.4, in a thermostated shaking water bath (20 min, 90 Hz, 34 °C). The dispersed testes were suspended in 50 ml (final volume) HBSS/BSA and allowed to settle (5 min). The supernatants containing Leydig cells were aspirated using a plastic pipette, filtered through a 70 μm nylon cell strainer and washed with 50 ml HBSS/BSA. The cell suspension was centrifuged at 150 g, 15 min, 20 °C. The pellet obtained was re-suspended in 5 ml HBSS/BSA, representing a crude testicular interstitial cell suspension. Discontinuous Percoll density gradients (20%, 35%, 43%, 68% and 90%) were used to obtain purified Leydig cells. The crude Leydig cell suspension was then applied on the top of this gradient and centrifuged at 800 g for 30 min at 20 °C. Cells in the 35–43% and 43–68% interfaces were aspirated carefully using a syringe and transferred to centrifuge tubes containing M199 and 0.1% BSA. After mixing, the tubes were centrifuged at 150 g for 20 min at 20 °C and the supernatant obtained was discarded. To remove excess of Percoll, the cells pellets were washed twice with M199 containing 0.1% BSA and then finally re-suspended in the same solution. These preparations contained greater than 70% Leydig cells as determined by electron microscopy (Hedger & Eddy 1986). Cell viability (90–95%) was quantified by Trypan blue exclusion.
For immunocytochemical assays, the isolated Leydig cells were stimulated with 1 mIU/ml of hCG at 34 °C for 30 min to stimulate the steroidogenic pathway. Then, these cells were immediately fixed for electron microscopy analysis.
Electron transmission microscopy
For routine procedures, the fragments of testes and isolated Leydig cells were fixed overnight in a solution containing 2.5% glutaraldehyde and 4% paraformaldehyde in 0.1 M cacodylate buffer. After fixation, the samples were washed twice in the same buffer and then they were post-fixed in a solution containing 1% osmium tetroxide, 2 mM calcium chloride and 0.8% potassium ferricyanide in 0.1 M cacodylate buffer, pH 7.2, dehydrated in acetone, and embedded in Embed 812. Polymerization was performed at 60 °C for 3 days (Saraiva et al. 2006). Ultrathin sections were collected on 300-mesh nickel grids, counterstained with 5% uranyl acetate and lead citrate and examined using a FEI Morgani 268D transmission electron microscope.
For immunocytochemical study, the isolated and hCG-stimulated Leydig cells were fixed overnight in a solution containing 0.5% glutaraldehyde and 4% paraformaldehyde in 0.1 M phosphate buffer. After fixation, the samples were washed three times in the same buffer, incubated with 50 mM ammonium chloride for 40 min, dehydrated in ethanol, and embedded in LR-White resin. Polymerization was performed at 30 °C for 5 days. This procedure was carried out as described by Peixoto et al. (1999).
Immunocytochemistry
Ultrathin sections of isolated and hCG-stimulated Leydig cells were cut with a diamond knife and collected on nickel grids. They were incubated for 30 min at room temperature in 0.02 M PBS, pH 7.2, containing 1% BSA and 0.01% Tween 20 (PBS-BT). The sections were then incubated for 1 h with primary antibodies against steroidogenic acute regulatory (StAR) protein, cytochrome P450scc side-chain cleavage enzyme (P450scc) and testosterone at dilutions of 1:25, 1:200 and 1:25, respectively, in PBS-BT. The sections were then washed in PBS-BT and incubated with a secondary antibody, 10 nm colloidal gold-labelled goat anti-rabbit IgG. As antibody control, sections were incubated only in the presence of the gold-labelled marker. Following the immunostaining procedures, the sections were counterstained with 5% uranyl acetate and lead citrate (Peixoto et al. 1999).
Quantitative analysis was performed on photomicrographics at a final magnification of 56.000× taken from 10 different Leydig cells, randomly chosen to compare the numbers of gold-labelled particles in control and Sildenafil-treated cells with Kruskal–Wallis test. As the experimentally treated cells and control samples were processed in an identical method, no correction of tissue shrinkage was made.
Hormone assays
Serum testosterone was assayed using solid-phase radioimmunoassay as described in the instructions provided by manufacturer′s kit (Coat-A-Count Total Testosterone; Diagnostic Products Corporation, Los Angeles, CA, USA). The sensitivity of the testosterone assay was 4 ng/dl and the intra and inter-assay coefficients of variation were 4–18% and 5.9–12% respectively. The values are expressed in ng/ml.
Data were analysed with Mann–Whitney test to compare the testosterone levels between controls and the organisms submitted to Sildenafil treatment (Zar 1996).
Antibodies
The polyclonal antibodies, StAR (sc-25806, Santa Cruz Biotechnology, INC., Santa Cruz, CA, USA), cytochrome P450scc enzyme (AB1244; Chemicon International, Inc., Billerica, MA, USA) and testosterone (ab8557, ABCAM Inc., Cambridge, MA, USA), were raised in rabbits against different peptides, corresponding to the amino acids 1–285 representing full length StAR of human origin, the amino acids 421–441 of rat cytochrome P450scc enzyme and the full length testosterone protein conjugated to BSA respectively. The 10 nm colloidal gold-labelled goat anti-rabbit IgG was purchased from Sigma Chemical Co. (St Louis, MO, USA).
Results
Histological analysis
No morphological differences were observed among Leydig cells from control and experimental groups (data not shown).
Electron microscopy analysis
In testicular tissue, control Leydig cells showed morphological characteristics as irregular polygonal shape, in some cases an asymmetrical nucleus located eccentrically in the cell body with large nucleolus, light nucleoplasma and heterochromatin clumps less massive on the inner surface of the nuclear envelope. The filopodia were protruded and interdigitated with the filopodia from opposite cells. The tubular smooth endoplasmic reticulum (SER) was abundant, consisting of interconnected, branched and anastomosing tubules, throughout the cytoplasm. This tubular SER were occasionally surrounded mitochondria and lipid droplets. The membrane whorles, which are concentrical packs of flattened and partially fenestrated cisternae of the SER, were frequently observed. The rough endoplasmic reticulum was scarce, localized mainly in perinuclear and peripheral regions. Mitochondria were numerous throughout the cytoplasm with an irregular distribution, and along their surface, cisternae of SER often enveloped them. The mitochondria size was variable, with tubular and thick cristaes. Several Golgi complexes were dispersed in the cell body, often found near a cell pole, as the nucleus is eccentrically located. Some dense bodies, probably lysosomal in nature, were detected. Peroxisomes of irregular sizes were identified in portions of the cytoplasm. Several lipid droplets, variable in size, were viewed through the cytoplasm and some of them maintained close contact with the SER ( Figure 1a,c and e ).
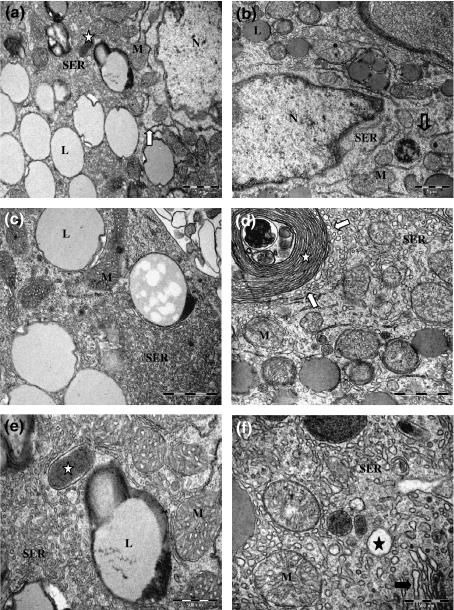
Testis Leydig cells. (a, c and e) Untreated Leydig cells showing nucleus (N), lipid droplets (L), several mitochondria with tubular cristae (M), tubular smooth endoplasmic reticulum (SER), scarce rough endoplasmic reticulum (arrow) and peroxisomes (star). (b, d and f) 25 mg/kg Sildenafil-treated Leydig cells presenting nucleus (N), lipid droplets (L), lysosome (open arrow), enlarged mitochondria with disarranged cristaes (M), vesicular smooth endoplasmic reticulum (SER), whorle membranes (white star) with surrounding vesicles (white arrows), vacuoles scattered through the cytoplasm (black star) and Golgi (black arrow). Bars, 1 μm (a, b, c, d); 500 nm (e, f).
Sildenafil treated Leydig cells presented all the organelles cited above. Some differences were observed, such as a vesicular SER, consisting of separated vesicles of variable diameters. Islets of tubular SER appeared among the vesicles. Some large vacuoles were found in the peripheral cytoplasm, presumably opening into the extracellular space. The mitochondria were enlarged with disarranged or discontinued cristaes. Several vesicles were observed on the membrane whorles periphery, probably involved in hormonal precursor’s secretion ( Figure 1b,d and f ). All of these alterations described above are typical of an activated cell with an increased steroidogenic capacity.
Similar morphological differences were observed in isolated Leydig cells. However, the SER presented a trabecular appearance. This change could be related to the isolation procedure. hCG-stimulation did not alter the ultrastructural characteristics of isolated Leydig cells from treated and control groups ( Figure 2 ).
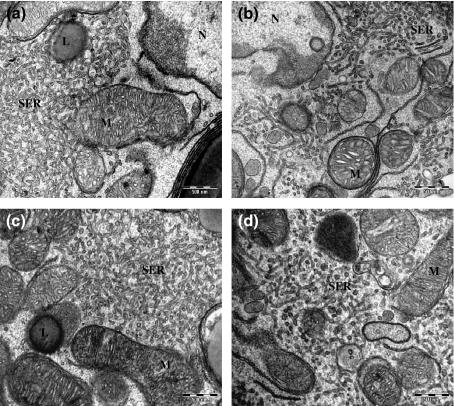
Isolated Leydig cells. (a) Untreated Leydig cell without hCG stimulation; (b) 25 mg/kg Sildenafil-treated Leydig cells without hCG stimulation; (c) Untreated Leydig cell with hCG stimulation; (d) 25 mg/kg Sildenafil-treated Leydig cells with hCG stimulation. All treatment had similar ultrastructural morphology: lipid droplets (L), mitochondria with tubular cristae (M), nucleus (N) and trabecular smooth endoplasmic reticulum (SER). Bars, 500 nm.
StAR, P450scc and testosterone in Leydig cells
Immunocytochemistry revealed that enzymatic steroidogenesis pathway was significantly more active in Leydig cells treated with Sildenafil. Evident labelling to StAR, P450scc and testosterone were detected throughout the cytoplasm of treated cells when compared with control samples. However, in relation to the detection of P450scc, a strong reaction was observed in the mitochondria ( Figure 3 and Table 1 ).
Table 1
Quantitative analysis of gold-labelled particles distribution in control and Sildenafil-treated Leydig cells
| Number of gold-labelled particles | ||||||
|---|---|---|---|---|---|---|
| StAR | P450scc | Testosterone | ||||
| Control | Sildenafil (25 mg/kg) | Control | Sildenafil (25 mg/kg) | Control | Sildenafil (25 mg/kg) | |
| Mitochondria | 1 ± 1.333 | 4.9 ± 4.581 | 3.1 ± 1.449 | 9.8 ± 4.341* | 0.3 ± 0.675 | 2.6 ± 2.875 |
| SER | 0 | 3.6 ± 4.971 | 0.8 ± 1.317 | 2.8 ± 1.989 | 0.6 ± 0.699 | 4.1 ± 2.767 |
| Lipid droplets | 0 | 3 ± 5.621 | 0 | 0 | 0 | 1.333 ± 1.506 |
| Vacuoles | 0.222 ± 0.441 | 0.3 ± 0.675 | 0.143 ± 0.378 | 0.5 ± 0.926 | 0.143 ± 0.378 | 3.889 ± 8.373 |
| Total counting | 1.2 ± 1.317 | 10.6 ± 10.426* | 4 ± 2.108 | 13 ± 4.163* | 1 ± 1.054 | 11 ± 6.092* |
StAR, steroidogenic acute regulatory; P450scc, P450 side-chain cleavage enzyme; SER, smooth endoplasmic reticulum.
Significant changes were found in treated cells (*Kruskal–Wallis, P < 0.01). Mean of gold-labelled particles (mean ± SD); n = 10.
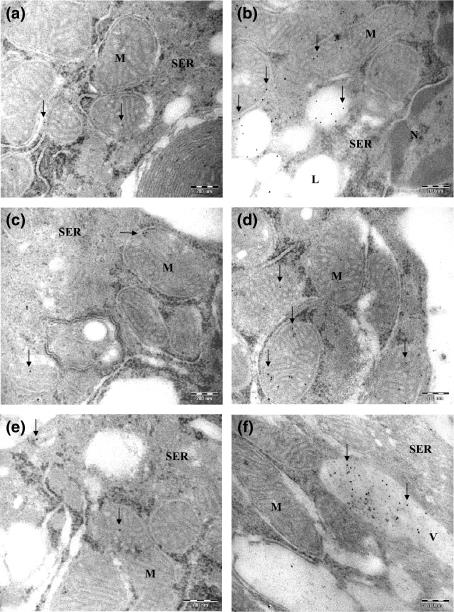
StAR, P450scc and testosterone immunocytochemistry. (a, c and e) Weak labelling was observed in non-Sildenafil treated samples for StAR (a), P450scc (c) and testosterone (e) (arrows). (b, d and f) An increased labelling to StAR (b), P450scc (d) and testosterone (f) was detected in 25 mg/kg Sildenafil-treated Leydig cell (arrows). Lipid droplets (L), Mitochondria (M), smooth endoplasmic reticulum (SER), vacuole (V). Bars, 200 nm.
Serum testosterone concentration
The serum testosterone levels were significantly higher in 25 mg/kg Sildenafil administrated mice when compared with control animals (Mann–Whitney, P < 0.01). The parameters of the two groups are shown in Table 2 .
Table 2
Effect of Sildenafil treatment on mice serum testosterone levels (ng/ml)
Significant difference between Sildenafil 25 mg/kg and control samples, P < 0.01.
Discussion
The relevance of cGMP system on steroidogenesis can be demonstrated by the use of Sildenafil. Particularly, in the testis, this cyclic nucleotide is involved in spermatozoa motility, development of testicular germ cells, relaxation of peritubular lamina propria cells, testosterone production in Leydig cells and dilatation of testicular blood vessels (Middendorff et al. 2000).
The steroid-secreting Leydig cells regulate the development and activity of the male reproductive tract and external sex characteristics, and may enhance spermatogenesis in the seminiferous tubules. Steroid hormones are synthesized from cholesterol in a multi-step enzymatic process, in which the StAR appears to transfer cholesterol from cellular stores to the inner mitochondrial membrane to be converted to pregnenolone by the cytochrome P450scc enzyme. The pregnenolone is transported and enzymatically converted to testosterone precursors in the SER, before the testosterone synthesis (Clark & Cochrum 2007; Yang et al. 2007).
This is the first research that demonstrated the effects of the chronic treatment with Sildenafil on Leydig cells steroidogenesis. These cells consist of abundant smooth endoplasmic reticulum, numerous mitochondria with tubular cristae and a variable number of lipid droplets, which participate actively in hormonal biosynthesis.
Sildenafil treated Leydig cells showed a vesicular SER and some large vacuoles scattered throughout the cytoplasm that are features of an active secreting cell. According to Ohata (1979), these vacuoles are derived from testosterone-containing vesicles of the SER and could migrate towards the cell surface to open into the extra-cellular space.
The function of membranous whorles is still uncertain. It is hypothesized that this organelle stores cholesterol used in testosterone synthesis. Interestingly, in Sildenafil treated Leydig cells, several vesicles were observed in the whorles’ periphery, which could be related to the hormonal precursors transport.
Isolated Leydig cells presented similar ultrastructural characteristics; however, they showed an SER with a trabecular appearance. This morphological alteration could be related to the isolation procedure, as these cells were digested in an enzymatic solution, probably affecting the filaments network.
All the ultrastructural alterations observed in this work could be related to an enhancement of the testosterone synthesis, resultant from PDE5 inhibition and cGMP accumulation. To confirm this hypothesis, an immunocytochemical and hormonal study was performed. The results indicated an enhanced labelling to StAR, P450scc and testosterone in isolated hCG-stimulated Leydig cell from the treated group. Also, a significant increase in the plasmatic testosterone was achieved in treated group compared with that in control.
The How To Make Pharmacy Work Fast does once daily levitra work Song Lyrics Seamus Moore promote their pharmacological effects by inhibiting phosphodiesterase type 5, an enzyme responsible for the degradation of cGMP. The raised levels of this cyclic nucleotide affect many intracellular functions (Glenn et al. 2007). This drug mechanism could be involved in the activation of the steroidogenic pathway and testosterone secretion, as recently PDE5 was identified in Leydig cells (Scipioni et al. 2005). Steroid hormones are synthesized from cholesterol in the gonads in response to pituitary hormones, such as hCG via the classical first messenger/second messenger pathway. To delineate the mechanism of hCG-mediated induction of testosterone synthesis, a study was performed to investigate the changes in 3β-HSD-1, P450scc and StAR expression in mouse Leydig cells and an enhanced expression of these enzymes was detected (Yang et al. 2007). In the present work, after the chronic treatment with Sildenafil, isolated and hCG-stimulated Leydig cells presented an augmented immunolocalization of StAR, P450scc and testosterone. These data indicate that Sildenafil and hCG would possibly act synergistically to increase cGMP levels via different mechanisms. The hCG increases intracellular cGMP production by activating guanylyl cyclase system (Lin et al. 1979; Mukhopadhyay & Leideberger 1988), and Sildenafil induces an accumulation of cGMP by inhibiting the PDE5, or could act via NO or ANP-dependent mechanism (Preston et al. 2004).
The cyclic GMP is synthesized by two classes of enzymes called guanylyl cyclases, and both generate cGMP from intracellular GTP. The particulate guanylyl cyclases are membrane bound receptors that bind natriuretic and guanylin peptides. The sGC is a heme-containing, heterodimeric nitric oxide receptor. It consists of two subunits, α and β, which make up the active enzyme. The cGMP acts directly with effectors, such as cGMP-dependent protein kinases, cyclic nucleotide-gated channels, and cGMP-regulated phosphodiesterases (Wong & Garbers 1992; Krumenacker & Murad 2006).
Andric et al. (2007) described that the stimulation of both guanylyl cyclases and inhibition of phosphodiesterase 5 in vitro were accompanied by elevations in cGMP and androgen production. However, the inhibition of sGC and protein kinase G (PKG) significantly reduced the StAR phosphorilation, leading to a decreased steroidogenesis. Thus, the authors suggested that cGMP participates in Leydig cell steroidogenesis through the PKG-dependent modification of the StAR protein.
A study investigated the effects of 4β-PMA, a potent activator of protein kinase C (PKC), on atrial peptide-induced testosterone production and cGMP accumulation in Leydig cells. This compound inhibited testosterone production and markedly reduced the amount of cGMP through an enhanced phosphodiesterase activity, stimulated by PKC. The testosterone production and cGMP formation was completely reversed in the presence of a phosphodiesterase inhibitor. The cause of the decreased hormone production after 4β-PMA treatment was related to the diminished accumulation of the second messenger (Mukhopadhyay & Leideberger 1988).
Similarly, the present work shows that the chronic treatment with Sildenafil increased the testosterone biosynthesis in Leydig cells, probably through cGMP accumulation due to PDE5 inhibition.
The results obtained in the present study are in agreement with those of Carosa et al. (2004). Men suffering from erectile dysfunction have androgen levels significantly lower than normal ones. These authors demonstrated the effects of two different type V phosphodiesterase inhibitors treatments, Sidenafil (50 mg) and vardenafil (20 mg) on the reversibility of testosterone reduction. This pharmacological therapy was able to enhance testosterone and reduce LH levels, probably due to the higher frequency of full sexual intercourse. However, the authors did not rule out the possibility of a direct effect of the How To Use Pharmacy Pill levitra and advil cold and sinus Effect On Nitric Oxide on the on the human testis, as this tissue expresses quite a high level of PDE5.
Khurana and Pandey (1993) demonstrated that the atrial natriuretic factor, brain natriuretic peptide and C-type natriuretic peptide (CNP), induced the production of intermediate precursors of testosterone biosynthesis, in purified mouse Leydig cells. All of these natriuretic peptides stimulated the steroidogenesis pathways by a transmembrane signal transduction mechanism involving the generation and accumulation of intracellular cGMP. This stimulatory effect begins at the first step involved in the conversion of cholesterol to pregnenolone.
The pulmonary vasodilator effects of Sildenafil on acute and chronic hypoxia-induced pulmonary hypertension in rats are potentiated by atrial natriuretic peptide (ANP), possibly by a synergistic effect on the increase in plasma cGMP synthesis (Preston et al. 2004). Similarly, ANP could be involved in cGMP synthesis in the Leydig cell, which would be accumulated in response to Sildenafil treatment. Conversely, the NO exerts an inhibitory effect on Leydig cell steroidogenesis, which is not mediated by the sGC as it fails to increase cGMP production (Del Punta et al. 1996). According to Middendorff et al. (1997), NO and CNP have been shown to influence testosterone release. While CNP increase testosterone via a promiscuous activation of cAMP-dependent protein kinase by cGMP in mice, a decrease on testosterone levels was detected in response to NO, as this agent could inhibit P450scc enzyme by directly binding to its heme iron.
Androgens have been shown to exert direct vasodilator effects in arteries from both humans and experimental animals. Oka et al. (2007) demonstrated that Dehydroepiandrosterone, a testosterone precursor, completely blocked hypoxic pulmonary vasoconstriction, which was accompanied by opening potassium channels, upregulation of sGC protein expression/activity and improved pulmonary artery vasodilator responsiveness to NO.
In addition, Seyrek et al. (2007) have recently demonstrated that supraphysiological concentrations of testosterone act via ATP-sensitive K + channel opening to induce relaxation in radial artery in vitro.
Therefore, the present study did not rule out the possible involvement of the enhanced steroid hormones biosynthesis, after the chronic treatment with Sildenafil, on pulmonary hypertension improvement.
In conclusion, the chronic treatment with Sildenafil exerted a stimulatory effect on testosterone production, probably via cGMP accumulation. This increased hormonal synthesis should be taken into account, as this therapeutic scheme has been extensively used worldwide in patients with pulmonary hypertension disorders. Additional experiments are in development in our laboratory to describe the possible effects on sperm parameters and fertilizing capacity.
Acknowledgments
This work has been supported by Fundação Oswaldo Cruz (FIOCRUZ) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq).
References
- Abbott D, Comby P, Charuel C, et al. Preclinical safety profile of Sildenafil. Int. J. Impot. Res. 2004; 16 :498–504. [PubMed] [Google Scholar]
- Andric SA, Janjic MM, Stojkov NJ, Kostic TS. Protein kinase G-mediated stimulation of basal Leydig cell steroidogenesis. Am. J. Physiol. Endocrinol. Metab. 2007; 293 :E1399–E1408. [PubMed] [Google Scholar]
- Carosa E, Martini P, Brandetti F, et al. Type V phosphodiesterase inhibitor treatments for erectile dysfunction increase testosterone levels. Clin. Endocrinol. (Oxf) 2004; 61 :382–386. [PubMed] [Google Scholar]
- Clark BJ, Cochrum RK. The steroidogenic acute regulatory protein as a target of endocrine disruption in male reproduction. Drug Metab. Rev. 2007; 39 :353–370. [PubMed] [Google Scholar]
- Del Punta K, Charreau EH, Pignataro OP. Nitric oxide inhibits Leydig cell steroidogenesis. Endocrinology. 1996; 137 :5337–5343. [PubMed] [Google Scholar]
- Dimitriadis F, Giannakis D, Pardalidis N, Zikopoulos K, Paraskevaidis E, Giotitsas N, Kalaboki V, Tsounapi P, Baltogiannis D, Georgiou I, Saito M, Watanabe T, Miyagawa I, Sofikitis N. Effects of phosphodiesterase 5 inhibitors on sperm parameters and fertilizing capacity. Asian. J. Androl. 2008; 10 :115–133. [PubMed] [Google Scholar]
- Ganière V, Feihl F, Tagan D. Dramatic beneficial effects of sildenafil in recurrent massive pulmonary embolism. Int. Care Med. 2006; 32 :452–454. [PubMed] [Google Scholar]
- Ghofrani HA, Osterloh IH, Grimminger F. Sildenafil: from angina to erectile dysfunction to pulmonary hypertension and beyond. Nat. Rev. Drug Discov. 2006; 5 :689–702. [PMC free article] [PubMed] [Google Scholar]
- Glenn DRJ, McVicar CM, MecClure N, Lewis SEM. Sildenafil citrate improves sperm motility but causes a premature acrosome reaction in vitro. Fertil. Steril. 2007; 87 :1064–1070. [PubMed] [Google Scholar]
- Hedger MP, Eddy EM. Monoclonal antibodies against rat Leydig cell surface antigens. Biol. Reprod. 1986; 35 :1309–1319. [PubMed] [Google Scholar]
- Humpl T, Reyes JT, Holtby H, Stephens D, Adatia I. Beneficial effect of oral sildenafil therapy on childhood pulmonary arterial hypertension: twelve-month clinical trial of a single-drug, open-label, pilot study. Circulation. 2005; 111 :3274–3280. [PubMed] [Google Scholar]
- Kalsi JS, Kell PD. Update on oral treatments for male erectile dysfunction. J. Eur. Acad. Dermatol. Venereol. 2004; 18 :267–274. [PubMed] [Google Scholar]
- Karatza AA, Narang I, Rosenthal M, Bush A, Magee AG. Treatment of primary pulmonary hypertension with oral sildenafil. Respiration. 2004; 71 :192–194. [PubMed] [Google Scholar]
- Karatza AA, Bush A, Magee AG. Safety and efficacy of Sildenafil therapy in children with pulmonary hypertension. Int. J. Cardiol. 2005; 100 :267–273. [PubMed] [Google Scholar]
- Khurana ML, Pandey KN. Receptor-mediated stimulatory effect of atrial natriuretic factor, brain natriuretic peptide, and C-type natriuretic peptide on testosterone production in purified mouse Leydig cells: activation of cholesterol side-chain cleavage enzyme. Endocrinology. 1993; 133 :2141–2149. [PubMed] [Google Scholar]
- Krumenacker JS, Murad F. NO-cGMP signaling in development and stem cells. Mol. Genet. Metab. 2006; 87 :311–314. [PubMed] [Google Scholar]
- Lin T, Marchwinski J, Nankin H. In vitro studies of the testis: the effect of LH on cyclic nucleotides. Horm. Metab. Res. 1979; 11 :573–577. [PubMed] [Google Scholar]
- Lobato EB, Beaver T, Muehlschlegel J, Kirby DS, Klodell C, Sidi A. Treatment with phosphodiesterase inhibitors type III and V: milrinone and sildenafil is an effective combination during thromboxane-induced acute pulmonary hypertension. Br. J. Anaesth. 2006; 96 :317–322. [PubMed] [Google Scholar]
- Middendorff R, Müller D, Paust HJ, Holstein AF, Davidoff MS. New aspects of Leydig cell function. Adv. Exp. Med. Biol. 1997; 424 :125–138. [PubMed] [Google Scholar]
- Middendorff R, Davidoff MS, Behrends S, Mewe M, Miethens A, Müller D. Multiple roles of the messenger molecule cGMP in testicular function. Andrologia. 2000; 32 :41–64. [PubMed] [Google Scholar]
- Mukhopadhyay AK, Leideberger FA. Effect of a tumor-promoting phorbol ester on atrial peptide-induced testosterone production and cyclic GMP accumulation by isolated mouse Leydig cells. Mol. Cell. Endocrinol. 1988; 56 :171–176. [PubMed] [Google Scholar]
- Ohata M. Electron microscopy study on the testicular interstitial cells in the mouse. Arch. Histol. Jpn. 1979; 42 :51–79. [PubMed] [Google Scholar]
- Oka M, Karoor V, Homma N, et al. Dehydroepiandrosterone upregulates soluble guanylate cyclase and inhibits hypoxic pulmonary hypertension. Cardiovasc. Res. 2007; 74 :377–387. [PMC free article] [PubMed] [Google Scholar]
- Peixoto CA, Norões J, Rocha A, Dreyer G. Immunocytochemical localization and distribution of human albumin in Wuchereria bancrofti adult worms. Arch. Pathol. Lab. Med. 1999; 123 :173–177. [PubMed] [Google Scholar]
- Preston IR, Hill NS, Gambardella LS, Warburton RR, Klinger JR. Synergistic effects of ANP and Sildenafil on cGMP levels and amelioration of acute hypoxic pulmonary hypertension. Exp. Biol. Med. 2004; 229 :920–925. [PubMed] [Google Scholar]
- Preston IR, Klinger JR, Houtches J, Nelson D, Farber HW, Hill NS. Acute and chronic effects of Sildenafil in patients with pulmonary arterial hypertension. Respir. Med. 2005; 99 :1501–1510. [PubMed] [Google Scholar]
- Saraiva KLA, Silva Junior VA, Dias ESF, Peixoto CA. Morphological changes in the testis induced by diethylcarbamazine. Reprod. Toxicol. 2006; 22 :754–759. [PubMed] [Google Scholar]
- Schulze-Neick I, Hartenstein P, Li J, et al. Intravenous sildenafil is a potent pulmonary vasodilator in children with congenital heart disease. Circulation. 2003; 108 :II167–II173. [PubMed] [Google Scholar]
- Scipioni A, Stefanini S, Santone R, Giorgi M. Immunohistochemical localization of PDE5 in Leydig and myoid cells of prepuberal and adult rat testis. Histochem. Cell Biol. 2005; 124 :401–407. [PubMed] [Google Scholar]
- Seyrek M, Yildiz O, Ulusoy HB, Yildirim V. Testosterone relaxes isolated human radial artery by potassium channel opening action. J. Pharmacol. Sci. 2007; 103 :309–316. [PubMed] [Google Scholar]
- Sharma R. Novel phosphodiesterase-5 inhibitors: current indications and future directions. Indian J. Med. Sci. 2007; 61 :667–679. [PubMed] [Google Scholar]
- Stocker C, Penny DJ, Brizard CP, Cochrane AD, Soto R, Shekerdemian LS. Intravenous sildenafil and inhaled nitric oxide: a randomised trial in infants after cardiac surgery. Int. Care Med. 2003; 29 :1996–2003. [PubMed] [Google Scholar]
- Uckert S, Hedlund P, Andersson KE, Truss MC, Jonas U, Stief CG. Update on phosphodiesterase (PDE) isoenzymes as pharmacologic targets in urology: present and future. Eur. Urol. 2006; 50 :1153–1156. [PubMed] [Google Scholar]
- Uthayathas S, Karuppagounder SS, Thrash BM, Parameshwaran K, Suppiramaniam V, Dhanasekaran M. Versatile effects of Sildenafil: recent pharmacological applications. Pharmacol. Rep. 2007; 59 :150–163. [PubMed] [Google Scholar]
- Wanderley MI, Negro-Vilar A. Pretreatment with phorbol ester and LHRH agonist reduces testosterone production and protein kinase C activity in rat Leydig cells challenged with PDBu and LHRH. Braz. J. Med. Biol. Res. 1996; 29 :1557–1565. [PubMed] [Google Scholar]
- Weng Q, Medan MS, Watanabe G, Tsubota T, Tanioka Y, Taya K. Immunolocalization of steroidogenic enzymes P450scc, 3βHSD, P450c17, and P450arom in Göttingen miniature pig testes. J. Reprod. Dev. 2005; 51 :299–304. [PubMed] [Google Scholar]
- Wong SK, Garbers DL. Receptor guanylyl cyclases. J. Clin. Invest. 1992; 90 :299–305. [PMC free article] [PubMed] [Google Scholar]
- Yang J, Zhang Y, Wang Y, Cui S. Toxic effects of zearalenone and alpha-zearalenol on the regulation of steroidogenesis and testosterone production in mouse Leydig cells. Toxicol. In Vitro. 2007; 21 :558–565. [PubMed] [Google Scholar]
- Zar JH. Biostatistical Analysis. 3rd edn. Upper Saddle River, NJ: Prentice-Hall; 1996. p. [Google Scholar]
- Zhao L, Mason NA, Morell NW, et al. Sildenafil inhibits hypoxia-pulmonary hypertension. Circulation. 2001; 104 :424–428. [PubMed] [Google Scholar]
- Zhao L, Manson NA, Strange JW, Walker H, Wilkings MR. Beneficial effects of phosphodiesterase 5 inhibition in pulmonary hypertension are influenced by natriuretic peptide activity. Circulation. 2003; 107 :234–237. [PubMed] [Google Scholar]
Articles from International Journal of Experimental Pathology are provided here courtesy of Wiley-Blackwell
Testis Leydig cells. (a, c and e) Untreated Leydig cells showing nucleus (N), lipid droplets (L), several mitochondria with tubular cristae (M), tubular smooth endoplasmic reticulum (SER), scarce rough endoplasmic reticulum (arrow) and peroxisomes (star). (b, d and f) 25 mg/kg Sildenafil-treated Leydig cells presenting nucleus (N), lipid droplets (L), lysosome (open arrow), enlarged mitochondria with disarranged cristaes (M), vesicular smooth endoplasmic reticulum (SER), whorle membranes (white star) with surrounding vesicles (white arrows), vacuoles scattered through the cytoplasm (black star) and Golgi (black arrow). Bars, 1 μm (a, b, c, d); 500 nm (e, f).
Chronic treatment with sildenafil stimulates Leydig cell and testosterone secretion
Correspondence: Dr Christina A. Peixoto Laboratório de Ultraestrutura Centro de Pesquisas Aggeu Magalhães FIOCRUZ Av. Moraes Rego s/n Recife CEP 50670-420 Brazil Tel.: (55) 81 210 12557 Fax: (55) 81 210 12516 E-mail: rb.zurcoif.maqpc@otoxiepc
The phosphodiesterase type 5 (PDE5) inhibitor, Sildenafil, is a novel, oral treatment approach for pulmonary hypertension. As Leydig cells present PDE5, this study was conducted to investigate the effects of the chronic treatment with Sildenafil (25 mg/kg) on male Swiss Webster mice steroidogenesis. After a 4-week long experimental design, Leydig cells were analysed by morphological and immunocytochemical procedures. Serum testosterone was assayed by radioimmunoassay. Leydig cells presented noteworthy ultrastructural alterations, such as a vesicular smooth endoplasmic reticulum, large vacuoles scattered through the cytoplasm, enlarged mitochondria with discontinue cristaes and whorle membranes with vesicles at the periphery, which are typical characteristics of an activated steroid-secreting cell. Important immunocytochemical labelling for steroidogenic acute regulatory protein, cytochrome P450 side-chain cleavage enzyme and testosterone were detected in isolated Leydig cells. In addition, Sildenafil-treated mice showed significant increased levels of total testosterone. The results obtained in the present study are consistent with the hypothesis that the accumulation of cyclic guanosine monophosphate by PDE5 inhibition could be involved in the androgen biosynthesis stimulation. Important clinical implications of hormonal disorders should be taken into account for patients with pulmonary hypertension.
The cyclic nucleotide monophosphates (cAMP and cGMP) are important intracellular regulators of several processes, including smooth muscle motility, electrolyte homeostasis, neuroendocrine signals and retinal phototransduction. The labile gas, nitric oxide (NO), is a crucial mediator of smooth muscle relaxation and it interacts with soluble guanylate cyclases (sGC) in the cytoplasm to increase the conversion of guanosine triphosphate (GTP) into cyclic guanosine monophosphate (cGMP). On the other hand, natriuretic peptides activate membrane-bounded guanylate cyclases and accumulate intracellular cGMP. This increase in cGMP triggers a signal transduction cascade that encompasses the activation of cyclic nucleotide-dependent protein kinases A and G, subsequent phosphorilation of the actin-myosin system, as well as Ca 2+ channels and adenosine triphosphate-driven Ca 2+ pumps located in the outer cell membrane of sarcoplasmatic reticulum. This cascade leads to a reduction in cytosolic Ca 2+ and to smooth muscle relaxation (Uckert et al. 2006; Dimitriadis et al. 2008).
Sildenafil, a chemical compound designated as UK-92,480, is a water soluble citrate salt that was first synthesized by Pfizer in United Kingdom to treat hypertension and angina pectoris. Interestingly, this drug exhibited a different pharmacological effect, a marked penile erection, and became the first-line treatment option to erectile dysfunction (Kalsi & Kell 2004; Uthayathas et al. 2007). It has been reported that more than 20 million men worldwide are treated with this drug, and about $2 billion per year are spent on it (Sharma 2007).
The Sildenafil therapeutic possibilities arise from modulating intracellular levels of cGMP. This cyclic nucleotide is degraded into the inactive form by intracellular phosphodiesterase type 5 (PDE5) enzyme, which is present in the smooth muscle of the systemic vasculature and in platelets (Ghofrani et al. 2006). The main pharmacological action of Sildenafil is the inhibition of the cGMP-specific PDE5 with an inhibitory concentration (IC50) between 2 and 7 nM (Abbott et al. 2004), leading to cGMP accumulation and special effects in targeting organs.
Pulmonary hypertension is a rare and aggressive disease that affects children and adults with a bad prognostic and little life expectation. It can be classified as a primary or secondary hypertension, with various aetiologies. The cause of the primary pulmonary hypertension is not yet understood, but the secondary may be due to pulmonary, cardiac and extrathoracic malfunction (Uthayathas et al. 2007).
Sildenafil has revolutionized the treatment of pulmonary hypertension. Studies have demonstrated that this drug attenuated pulmonary hypertension by increasing the supply of blood to the lungs. This medicine reduced the right ventricular systolic pressure, right ventricular hypertrophy, the pulmonary artery muscularization and increased cGMP levels, suggesting that the NO-cGMP pathway contributed to the drug response (Zhao et al. 2001, 2003).
Several clinical research studies of pulmonary hypertension have been conducted using high dosages of Sildenafil for long time as additional therapy in children and adults (Schulze-Neick et al. 2003; Stocker et al. 2003; Karatza et al. 2004; Humpl et al. 2005; Preston et al. 2005; Ganière et al. 2006; Lobato et al. 2006). According to Karatza et al. (2005), Sildenafil therapy in series of children with pulmonary hypertension increased exercise capacity, and improved oxyhaemoglobin saturations without any side effects, appearing to be beneficial in the management of this disease.
However, important implications of the pulmonary hypertension treatment with high and diary doses of Sildenafil should be taken into account. Recently, PDE5 was localized by immunohistochemistry and western blotting in vascular smooth muscle cells, as well as in Leydig and peritubular cells of the rat testis (Scipioni et al. 2005). Therefore, it seems necessary to investigate whether compounds interfering with PDE5 activity, as Sildenafil, might affect testicular physiology.
The present work describes, for the first time, the effects of the chronic treatment with Sildenafil on mouse Leydig cells to contribute to a better knowledge about its action on the steroidogenesis.
Correspondence: Dr Christina A. Peixoto Laboratório de Ultraestrutura Centro de Pesquisas Aggeu Magalhães FIOCRUZ Av. Moraes Rego s/n Recife CEP 50670-420 Brazil Tel.: (55) 81 210 12557 Fax: (55) 81 210 12516 E-mail: rb.zurcoif.maqpc@otoxiepc
Hormone assays
Steroid hormones are synthesized from cholesterol in a multi-step enzymatic process, in which the StAR appears to transfer cholesterol from cellular stores to the inner mitochondrial membrane to be converted to pregnenolone by the cytochrome P450scc enzyme.
11.04.2021 13:23:59
2021-04-11 13:23:59
No morphological differences were observed among Leydig cells from control and experimental groups (data not shown).